
|
|
|
|
|
|
Shark Info (14.11.1997) |
Author |
|
Intro: |
Shark info |
|
Hauptartikel: |
Shark info |
|
Fact Sheet: |
Dr. J. G. New |
|
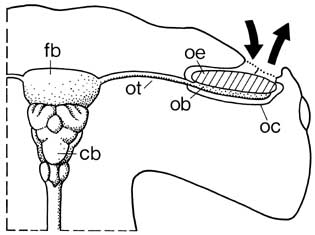
Fact Sheet: Der Geruchssinn des Hais«Die schwimmende Nase»Von Dr. John G. New Haie können Gerüche auch noch in milliardenfacher
Verdünnung aufspüren. Ihr gut entwickelter
Geruchssinn, die auffälligen, grossen Nasenlöcher und die damit verbundene Hirnregion
haben den Tieren auch die Bezeichnung schwimmende Nase eingetragen.
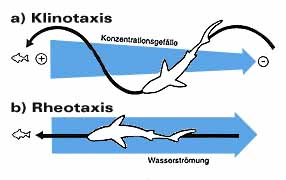
Physiologische Untersuchungen ergaben, dass der Geruchssinn dieser Meeresbewohner äusserst sensibel ist. So wurde nachgewiesen, dass Zitronenhaie (Negaprion brevirostris) beispielsweise eine 25-millionenfache Verdünnung von Thunfischextrakten entdecken und Schwarzspitzen-Riffhaie (Carcharhinus limbatus) und Graue Riffhaie (Carcharhinus amblyrhynchos) sogar Extrakte aus Muskelfleisch in einer Verdünnung von 1 zu 10 Milliarden orten. Das entspricht einem Tropfen in einem etwa tausend Quadratmeter grossen und zwei Meter tiefen Becken. Wie Haie ihre Beute mit dem Geruchssinn lokalisieren, ist noch nicht vollständig geklärt. Heute geht man von zwei Modellen aus: Eines dieser Modelle basiert auf der Tatsache, dass die Konzentration von Geruchsstoffen an deren Quelle am grössten ist und mit zunehmender Distanz abnimmt. Dieses Modell ( Gradient search model ) besagt, dass Haie die Konzentrationsveränderungen des Geruchsstoffes fortlaufend analysieren und sich von einer Umgebung mit schwächerer in eine solche mit stärkerer Konzentration begeben. Diese Suchform heisst Klinotaxis ; um sie zu optimieren, muss das Tier stets in einem bestimmten Winkel zur Geruchsquelle schwimmen und die Richtung ändern, sobald die Konzentration unter ein bestimmtes Mass sinkt. Diese Methode ergibt eine S-förmige Bewegung gegen die Quelle. Es gibt begründete Hinweise darauf, dass gewisse Haie diese Technik anwenden; obschon die Konzentrationsunterschiede in einigem Abstand zur Geruchsquelle dermassen gering sind, dass ein Lokalisieren schwierig ist. Ein zweites Modell beschreibt einen anderen Weg zur Geruchsquelle: Die Haie schwimmen nicht gegen das Konzentrationsgefälle, sondern gegen die Strömung ( Rheotaxis ). Bei diesem Modell ist das Tier gezwungen, andere sensorische Hilfen einzubeziehen wie etwa das Seitenlinienorgan, das die Strömungsrichtung erkennt. Dies wäre auch eine logische Erklärung, weil Geruchsstoffe im Wasser mit der Strömung ziehen. Haie scheinen die eine oder andere dieser Strategien oder auch beide zusammen zu benutzen. Vieles aus dem Geruchssensorik des Hais ist noch unbekannt. Zum Beispiel, ob chemische Gerüche auch zur Kommunikation verwendet werden, wie etwa das Anlocken von Geschlechtspartnern mit chemischen Lockstoffen. Die grosse Empfindlichkeit des Hai-Geruchssinns legt eine solche Vermutung nahe. Unbekannt ist zudem, ob Geruchshinweise beim Wanderverhalten eine Rolle spielen und wie die Informationen von Gerüchen im zentralen Nervensystem verarbeitet werden.
* Dr. John G. New ist Associate Professor im Department of Biology and Parmly Hearing Institute der Loyola Universität in Chicago und besitzt grosse Erfahrung im Forschungsbereich zum Octavolateralen System (Seitenlinien- und Innenohrsytem) ) der Haie.
Ausgewählte Literatur: Sheldon, R.E. (1911) The reaction of the dogfish to chemical stimuli. Journal of Comparative Neurology and Psychology 19: 723-311 Parker, G.H. (1914) The directive influence of the sense of smell in the dogfish. Bulletin of the U.S. Bureau of Fisheries 33:61-68. Tester, A.L. (1963) The role of olfaction in shark predation. Pacific Science 17: 145-170 Kleerekoper, H. (1978) Chemoreception and its interaction with flow and light perception in the locomotiona and orientation of some elasmobranchs. In: Sensory Biology of Sharks, Skates and Rays, RF Mathewson and E.S. Hodgson, eds. Office of Naval Research, Arlington, VA. Veröffentlichung nur mit Quellenangabe: Shark Info / Dr. John G. New |
||||||||||||||||||||||||
|
|
|
|
|
|||||